칼륨 통로
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
칼륨 통로는 전기화학적 기울기에 따라 칼륨 이온을 선택적으로, 빠르게 전달하는 막 단백질이다. 이러한 통로는 세포의 휴지 전위를 설정하고 활동 전위를 조절하는 데 중요한 역할을 하며, 심장 박동 조절, 호르몬 분비 등 다양한 생리적 과정에 관여한다. 칼륨 통로는 칼슘 활성화, 내향 정류, 탠덤 기공 도메인, 전압 개폐의 4가지 주요 유형으로 분류되며, 각 유형은 특정 자극에 반응하여 칼륨 이온의 흐름을 조절한다. 칼륨 통로의 기능 이상은 심장 부정맥, 제2형 당뇨병 등 다양한 질병을 유발할 수 있으며, 덴드로톡신과 같은 독소에 의해 차단될 수 있다. 칼륨 통로의 구조는 4개의 단백질 소단위체로 이루어진 4량체이며, 선택성 필터와 소수성 영역, 중심 공동 등의 특징적인 구조를 통해 칼륨 이온의 선택적 투과를 가능하게 한다. 칼륨 통로의 게이팅과 불활성화는 이온 흐름을 조절하는 중요한 과정이며, 약리학적으로 차단제와 활성제가 개발되어 연구되고 있다. 또한, 무스카린 칼륨 통로는 심박수 조절에 중요한 역할을 하며, 칼륨 통로를 모티브로 한 예술 작품도 존재한다.
더 읽어볼만한 페이지
- 이온 통로 - 니코틴성 아세틸콜린 수용체
니코틴성 아세틸콜린 수용체는 아세틸콜린에 의해 활성화되어 이온을 통과시키는 리간드 개폐 이온 채널 막단백질로, 척추동물에서 근육형과 신경형으로 나뉘어 신경 전달에 중요한 역할을 하며, 활성화는 뉴런 탈분극을 유발하고 니코틴 의존증과 같은 질병 위험과도 연관된다. - 이온 통로 - 고리형 뉴클레오타이드 개폐 이온 통로
고리형 뉴클레오타이드 개폐 이온 통로는 cAMP 또는 cGMP에 의해 활성화되는 이온 채널로, 다양한 생물학적 시스템에서 생체 신호 전달, 시각, 후각 등 다양한 생리적 기능을 수행하며, 구조는 4개의 단백질 소단위체로 구성된다. - 전기생리학 - 바이오피드백
바이오피드백은 정밀 기기로 생리적 활동을 측정하여 사용자에게 피드백을 제공, 사고, 감정, 행동 변화를 유도하여 생리적 변화를 지원하며, 치료 및 성적 향상에 활용되는 과정이다. - 전기생리학 - 근전도 검사
근전도 검사는 근육과 신경의 기능 이상을 진단하기 위해 근육의 전기적 활동을 측정하는 검사로, 신경근 질환 진단, 운동 연구, 보툴리눔 독소 주입 보조 등 다양한 분야에서 활용되며 인터페이스 제어 기술로도 연구된다. - 내재성 막 단백질 - 광계
광계는 틸라코이드 막에 위치하며 빛 에너지를 화학 에너지로 전환하는 단백질 복합체로, 광수집 복합체에 둘러싸인 반응 중심을 가지며 물 분해를 통해 전자를 얻고 산소를 방출하는 광계 II와 NADPH를 생성하는 광계 I 두 가지 주요 유형으로 나뉜다. - 내재성 막 단백질 - 사이토크롬 b6f 복합체
사이토크롬 b6f 복합체는 광합성에서 광계 II와 광계 I 사이의 전자 및 에너지 전달을 매개하고 틸라코이드 막을 가로질러 양성자를 수송하여 ATP 합성을 촉진하는 단백질 복합체로서, 순환적 광인산화에도 중요한 역할을 하며 사이토크롬 bc1 복합체와 구조적 유사성을 갖는 8개의 소단위체로 구성된 이량체이다.
| 칼륨 통로 | |
|---|---|
| 기본 정보 | |
 | |
| 설명 | 칼륨 이온을 선택적으로 투과시키는 이온 채널 |
| 세부 사항 | |
| 구조 | 4개의 동일한 하위 단위 또는 4개의 단백질로 구성됨 각 하위 단위는 채널의 기공 루프를 구성하는 2개의 막 스패닝 도메인 사이에 기공 루프를 포함함 |
| 선택성 필터 | 채널의 가장 좁은 부분 칼륨 이온이 나트륨과 같은 다른 이온보다 훨씬 쉽게 통과할 수 있도록 설계됨 |
| 기능 | |
| 역할 | 세포막 전위 설정 활동 전위 형성 세포의 전기적 흥분성 조절 |
| 활성화 | 전압에 의해 리간드 결합에 의해 다른 자극에 의해 |
| 유형 | |
| 종류 | 전압 개폐 칼륨 채널 (Voltage-gated K+ channels, Kv) 칼슘 활성화 칼륨 채널 (Calcium-activated K+ channels) 내부 정류 칼륨 채널 (Inward rectifier K+ channels, Kir) 2-P 도메인 칼륨 채널 (Two-pore-domain potassium channels, K2P) |
| 추가 정보 | |
| 관련 질병 | 부정맥 뇌전증 당뇨병 |
| 연구 | 로드 맥키넌 칼륨 채널의 구조와 메커니즘에 대한 연구로 2003년 노벨 화학상 수상 |
2. 기능
칼륨 통로는 전기화학적 기울기를 따라 칼륨 이온(K+)을 빠르고 선택적으로 전달하는 기본적인 기능을 수행한다. 이 전달 속도는 물속에서 칼륨 이온이 확산 속도에 버금갈 정도로 빠르며, 이온 반경 차이가 1 Å 미만인 나트륨 이온은 효과적으로 배제할 정도로 높은 선택성을 보인다.[4]
생물학적으로 칼륨 통로는 다양한 세포 기능에 필수적인 역할을 한다.
- 신경계: 많은 세포에서 휴지 전위를 설정하거나 재설정하는 데 관여한다. 특히 뉴런과 같은 흥분성 세포에서는 칼륨 이온의 지연된 역류를 통해 활동 전위를 형성하고 조절한다.
- 순환계: 심근 세포에서는 심장 활동 전위의 지속 시간을 조절하는 데 기여한다. 만약 칼륨 통로 기능에 문제가 생기면 생명을 위협할 수 있는 심장 부정맥이나 심부전 등을 유발할 수 있다. 또한 혈관 긴장을 유지하는 데에도 관여할 수 있다.
- 내분비계: 호르몬 분비를 조절하는 과정에도 참여한다. 예를 들어, 췌장의 베타 세포에서 인슐린 방출을 조절하는 데 관여하며, 이 기능에 문제가 생기면 제2형 당뇨병과 같은 질병으로 이어질 수 있다.
덴드로톡신과 같은 일부 독소는 칼륨 통로를 차단하여 신경계 등에 심각한 영향을 미칠 수 있다.[5]
3. 종류
칼륨 통로에는 네 가지 주요 종류가 있다.
- 칼슘 활성화 칼륨 통로: 칼슘 이온이나 다른 신호 분자의 존재에 반응하여 열린다.
- 내향 정류 칼륨 이온 통로: 내향 방향(세포 안으로)으로 전류(양전하)를 더 쉽게 통과시킨다.
- 탠덤 기공 도메인 칼륨 통로: "휴지 칼륨 통로" 또는 뉴런의 음극 막 전위를 설정하는 "누설 통로"와 같이 구성적으로 열리거나 높은 기저 활성을 갖는다.
- 전압 개폐 칼륨 통로: 전압 개폐 이온 통로이며, 막 전위의 변화에 반응하여 열리거나 닫힌다.
다음 표는 주요 칼륨 통로 종류와 대표적인 예를 비교하여 보여준다.
4. 구조
칼륨 통로는 기본적으로 4개의 동일하거나[54] 유사한 단백질 소단위체가 모여 중앙에 이온 통로를 형성하는 4량체 구조를 가진다.[55] 이 소단위체들은 중앙의 이온 전도 통로를 중심으로 4중 대칭(C4) 배열을 이루는 동종사량체(homotetramer)를 형성하거나, 서로 관련된 4개의 소단위체가 유사한 C4 대칭성을 가진 이종사량체(heterotetramer) 복합체를 형성할 수도 있다. 모든 칼륨 통로 소단위체는 공통적으로 통로의 위쪽 부분을 형성하며 칼륨 이온에 대한 선택적 투과성을 결정하는 독특한 통로 루프(pore loop) 구조를 가지고 있다.
포유류에는 칼륨 통로 단백질 소단위체를 암호화하는 유전자가 80개 이상 존재한다. 하지만 분자 구조 연구는 주로 세균에서 발견되는 칼륨 통로를 모델로 활발히 진행되었다. 특히 방선균 유래의 KcsA는 X선 결정학을 이용하여 그 구조가 로드 매키넌 연구팀에 의해 처음 밝혀졌으며,[103] 이를 통해 칼륨 이온이 어떻게 이 통로를 통과하며, 크기가 더 작은 나트륨 이온은 통과하지 못하는지에 대한 중요한 정보들이 밝혀졌다.[56] 매키넌은 이러한 선구적인 연구 업적을 인정받아 2003년 노벨 화학상을 수상했다.[57] 이후 인간을 포함한 다양한 생물의 칼륨 통로 구조가 연구되고 있다.
칼륨 통로는 크게 두 부분으로 나눌 수 있다. 이온이 통과하는 길을 형성하는 포어 도메인(pore domain)과, 막 전위 변화나 리간드 결합 같은 외부 자극을 감지하는 자극 수용 도메인(stimulus-sensing domain)이다. 포어 도메인의 구조는 거의 모든 종류의 칼륨 통로에서 매우 유사하게 보존되어 있는 반면, 자극 수용 도메인은 통로의 종류에 따라 다양한 구조를 가진다.
포어 도메인은 4개의 소단위체가 대칭적으로 배열되어 형성된다. 각 소단위체는 두 개의 막관통 나선과 이 둘을 연결하는 포어 루프(P 루프)로 구성된다. 이 4량체의 중앙에는 이온이 통과하는 통로가 만들어지는데, 이 통로는 세포 바깥쪽 부분에 위치하여 이온 선택성을 결정하는 선택성 필터, 통로 중앙의 넓은 공간인 중심 공동, 그리고 세포 안쪽 부분에 위치하여 통로 개폐를 조절하는 나선형 게이트(helical gate)로 이루어져 있다.
4. 1. 선택성 필터 (Selectivity filter)
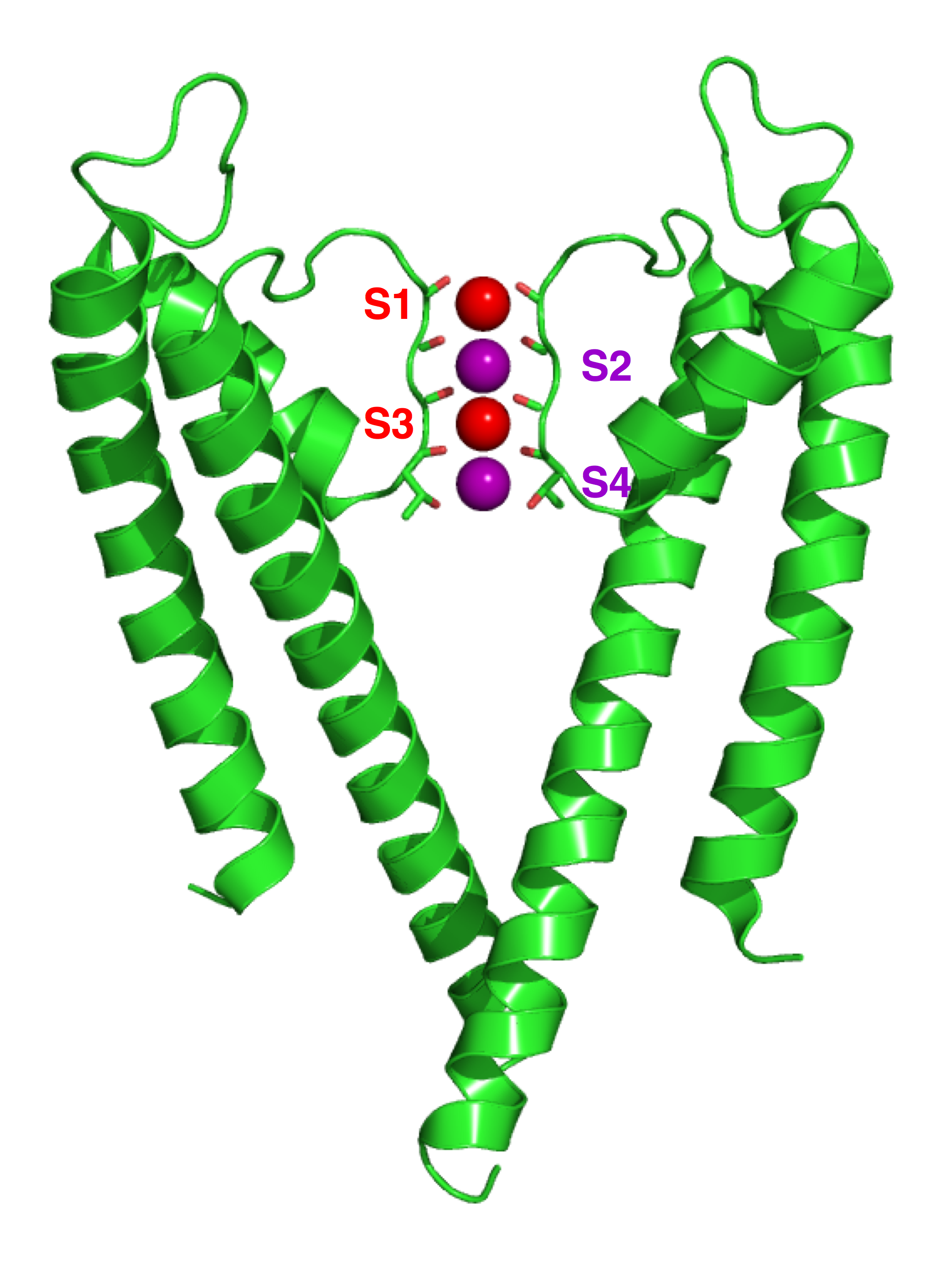
칼륨 이온은 통로의 선택성 필터(selectivity filter)에 들어갈 때 주변의 물 분자들로 이루어진 수화 껍질을 벗게 된다.[59] 이 선택성 필터는 칼륨 통로를 구성하는 4개의 소단위체 각각에 존재하는 특정 아미노산 잔기 서열에 의해 형성된다. 이 서열은 보통 원핵생물에서는 TVGYG (트레오닌-발린-글리신-타이로신-글리신), 진핵생물에서는 TIGYG (트레오닌-아이소류신-글리신-타이로신-글리신) 형태로 나타나며, 칼륨 통로의 '시그니처 시퀀스(signature sequence)'라고 불린다.[59] 이 시그니처 시퀀스는 P-루프(P-loop)라고 하는, 막 단백질의 특정 구조 부위 내에 위치한다.[59] 이 서열과 구조는 다양한 종류의 칼륨 통로에서 매우 유사하게 보존되어 있으며,[59] 단백질 구조 모티프 중 하나인 '둥지(nest)' 모티프와 유사한 독특한 3차원 구조를 이룬다.[59]
선택성 필터의 핵심 구조는 시그니처 시퀀스를 구성하는 아미노산들의 주쇄(backbone)에 있는 카르보닐 산소 원자들이 통로의 중심을 향해 정렬되어 있는 형태이다.[59] 이 4개의 전기 음성적인 카르보닐 산소 원자들이 각 칼륨 이온 결합 부위 주변에서, 마치 물 분자가 이온을 둘러싸는 것과 유사한 기하학적 구조(사각 반각기둥, square antiprism)를 형성한다.[59] 필터 내 카르보닐 산소와 칼륨 이온 사이의 거리는 수용액 상태에서 물 분자의 산소 원자와 칼륨 이온 사이의 거리와 거의 같기 때문에, 칼륨 이온은 수화 껍질을 벗고 필터 안으로 들어가는 과정에서 에너지적으로 큰 손실 없이 안정화될 수 있다.[59] 즉, 칼륨 이온은 탈수화된 상태에서도 필터 내 카르보닐 산소와의 상호작용을 통해 안정적으로 존재하며 통로를 통과할 수 있다. 선택성 필터 내에는 이렇게 칼륨 이온이 안정적으로 머무를 수 있는 결합 부위가 4곳 존재한다.
반면, 나트륨 이온은 칼륨 이온보다 크기가 작아서 필터 내 카르보닐 산소 원자들이 만드는 공간에 딱 맞게 들어가지 못한다.[59] 따라서 나트륨 이온은 탈수화되어 필터를 통과하는 것보다 세포 외부에서 물 분자와 결합된 상태로 남아있는 것이 에너지적으로 더 유리하기 때문에, 칼륨 통로를 잘 통과하지 못한다.[59] 이러한 구조적 특징이 칼륨 통로가 칼륨 이온을 선택적으로 통과시키는 중요한 원리이다.
선택성 필터의 정교한 구조는 필터를 둘러싸고 있는 방향족 아미노산 잔기들 사이의 수소 결합과 반 데르 발스 힘에 의해 안정적으로 유지되는 것으로 여겨진다.[54][60]
칼륨 통로의 이러한 구조와 기능은 방선균 유래의 KcsA 칼륨 통로의 결정 구조가 로데릭 매키넌 연구팀에 의해 처음 밝혀지면서 구체적으로 이해되기 시작했다.[103] 이후 인간을 포함한 다양한 생물의 칼륨 통로 구조가 연구되고 있다.
4. 1. 1. 선택성 기전 (Selectivity mechanism)
칼륨 이온 통로의 선택성 기전은 지속적인 논쟁의 대상이다. 통로는 이온이 선택적 필터에 들어갈 때 수화 껍질을 제거한다.[59] 이 선택적 필터는 각 4개의 소단위체 내에 위치한 시그니처 시퀀스(signature sequence)인 TVGYG (원핵생물) 또는 TIGYG (진핵생물)라는 5개의 아미노산 잔기 서열로 형성된다. 이 서열은 P-루프(P-loop)라고 불리는, 기공 헬릭스와 막관통 영역(TM2/6) 사이의 루프 안에 있으며, 칼륨 통로 간에 매우 잘 보존되어 있다.[59] 이 시퀀스는 단백질 구조 모티프 중 '둥지(nest)' 모티프와 구조적으로 유사한 독특한 주쇄 구조를 채택한다.[59]
선택 필터 내에서는 각 아미노산의 주쇄 카르보닐 산소 원자가 이온 투과 경로 쪽으로 향하고 있다. 4개의 전기 음성적인 카르보닐 산소 원자 세트가 필터 기공 중심을 향해 정렬되어, 각 칼륨 이온 결합 부위 주위에 물을 용매화시키는 껍질과 유사한 사각 반각기둥(square antiprism) 구조를 형성한다.[59] 필터 내 카르보닐 산소와 칼륨 이온 사이의 거리는 수용액에서 첫 번째 수화 껍질의 물 산소와 칼륨 이온 사이의 거리와 동일하여, 이온이 탈수화되는 데 유리한 에너지 경로를 제공한다.[59] 이처럼 칼륨 이온은 탈수화된 상태에서도 카르보닐 산소와의 배위를 통해 안정적으로 존재할 수 있다. 선택 필터의 구조적 안정성은 필터를 둘러싸는 방향족 아미노산 잔기 시트 내의 수소 결합과 반 데르 발스 힘에 의해 유지되는 것으로 보인다.[54][60]
반면, 나트륨 이온은 크기가 작아 카르보닐 산소 원자 사이의 공간을 효과적으로 채우지 못한다. 이 때문에 나트륨 이온은 칼륨 선택적 기공을 통과하는 것보다 세포 외부 공간에서 물 분자와 결합된 상태를 유지하는 것이 에너지적으로 더 유리하여 통과하지 못한다.[59] 칼륨 이온의 선택성은 이러한 구조적 특징과 더불어, 통로와 각 이온의 친화성 차이, 그리고 선택 필터 내에 4개의 칼륨 이온 결합 부위가 일렬로 배열된 점 때문에 발생하는 것으로 제안된다.
선택 필터는 일반적으로 세포 외부에서 시작하여 S1에서 S4까지 4개의 위치에서 칼륨 이온을 수용할 수 있다. 또한, 이온은 단백질 중앙의 물로 채워진 공동(cavity) 내 SC 위치나, 세포 외부의 S0 또는 Sext라고 불리는 위치에도 결합할 수 있다. 이러한 여러 위치에 이온이 동시에 점유하는 것이 가능하지만, 인접한 두 위치를 동시에 점유하는 것은 정전기적 반발 때문에 에너지적으로 불리하다. X선 구조는 많은 분자의 평균 상태를 보여주므로, 실제 점유 상태를 직접 추론하기는 어렵다.
KcsA의 이온 이동 기전은 이론적 계산과 시뮬레이션을 통해 광범위하게 연구되었다.[67][68] 연구 결과, 두 개의 이중 점유 상태 (S1, S3)와 (S2, S4)가 이온 전도에 필수적인 역할을 하는 것으로 나타났다. 분자 역학(MD) 시뮬레이션은 또한 이온이 필터에 들어가고 나가는 것을 반영하는 두 개의 세포외 상태, Sext 및 S0가 이온 전도에 중요하다고 제안한다.
이온 투과 경로의 중앙 부근에는 약 10 Å 정도 넓이의 중심공(central cavity)이 있다. 세포 내부에서 외부로 칼륨 이온이 이동할 때, 중심공까지는 수화된 상태로 도달하고, 중심공에서 선택 필터로 전달될 때 탈수화된다. P 루프 안쪽 헬릭스의 C 말단 쪽이 중심공을 향하고 있으며, 이 헬릭스의 전기 쌍극자 모멘트 때문에 중심공은 약간 음전하를 띠게 된다. 이로 인해 중심공에서 칼륨 이온과 같은 양이온이 안정적으로 존재할 수 있게 된다.
선택성 기전에 대한 이해는 분자 역학 시뮬레이션,[61] 이온 결합 모형,[62] 열역학적 계산,[63] 위상학적 고려,[64][65] 그리고 선택적 통로와 비선택적 통로 간의 구조적 차이 비교[66] 등 다양한 연구 방법을 통해 발전해왔다.
4. 2. 소수성 영역 (Hydrophobic region)
이 영역은 칼륨 이온 주위의 환경을 중화시켜 어떤 전하에도 끌리지 않게 한다. 결과적으로 반응 속도를 높인다.4. 3. 중심 공동 (Central cavity)
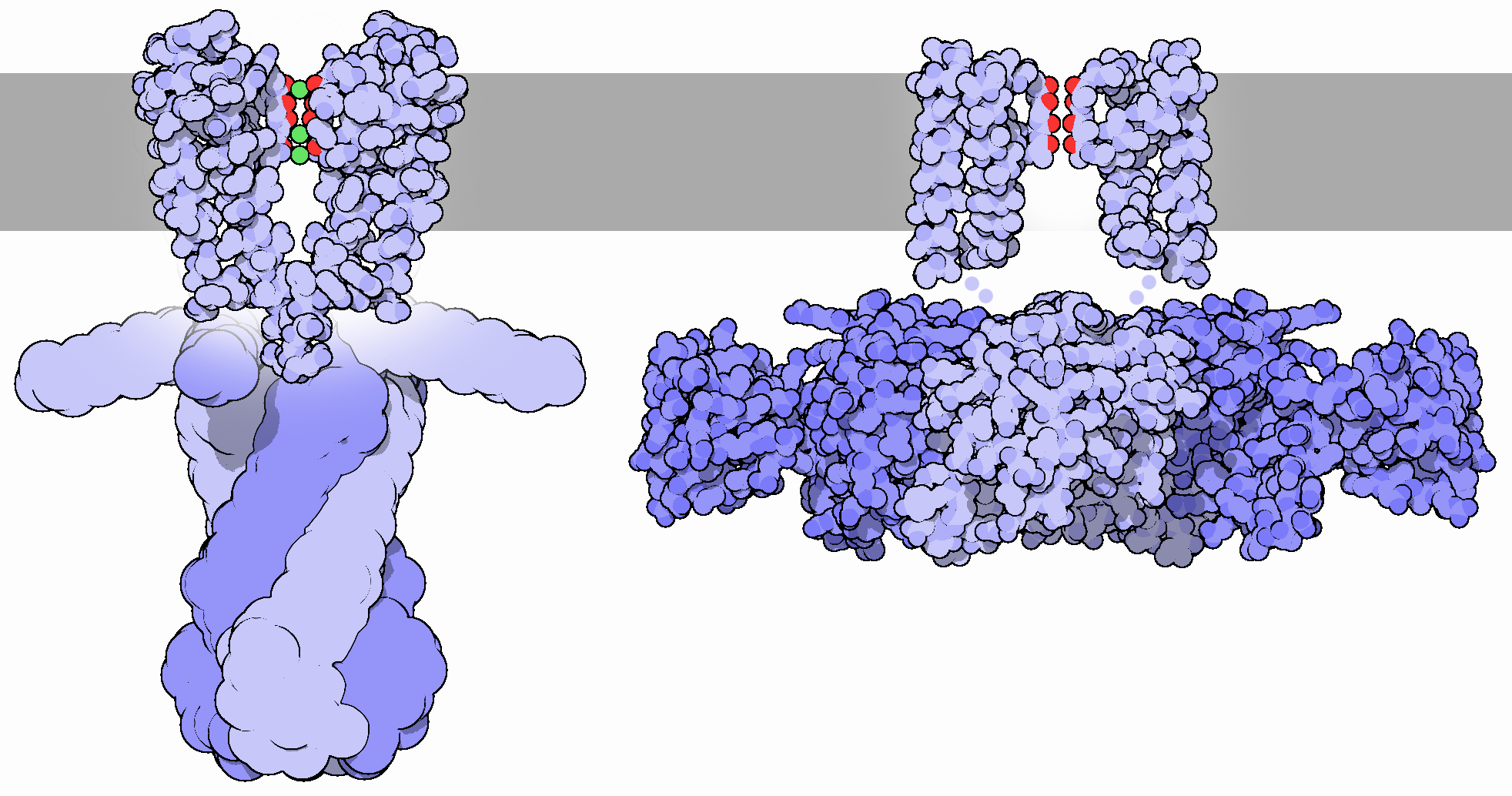
중심 공동(Central cavity)은 칼륨 통로의 이온 투과 경로 중앙 부근에 위치한 공간으로, 그 너비는 약 10 옹스트롬 정도이다.[103] 이 영역은 채널 벽의 소수성 특성으로 인해 통과하는 이온에 대한 에너지 장벽이 가장 높다.thumb은 빨간색과 파란색 선으로 표시되어 있다. 보라색 구는 칼륨 이온을 나타낸다.]]
그러나 중심 공동은 물 분자로 채워져 있으며, 이온 투과 경로를 구성하는 포어 루프(P 루프) 내 헬릭스의 극성 C 말단과 함께 이온이 넘어야 할 에너지 장벽을 낮추는 역할을 한다. P 루프 헬릭스의 C 말단은 중심 공동을 향해 배열되어 있는데, 이 헬릭스가 가지는 전기 쌍극자 모멘트의 영향으로 중심 공동 내부는 약간의 음전하를 띠게 된다. 이러한 환경은 칼륨 이온(K+)과 같은 양이온이 안정적으로 머무를 수 있도록 돕는다.
또한, 중심 공동의 존재는 칼륨 이온을 물 분자가 많은 고유전율 환경에 머무르게 함으로써, 세포막과 같이 유전율이 낮은 환경이 만드는 전기적 반발력(유전 장벽)을 극복하는 데 중요한 역할을 한다.
세포 내부에서 수화된 상태로 이동해 온 칼륨 이온은 중심 공동까지 도달한 후, 이곳에서 선택성 필터(selectivity filter)로 이동하는 과정에서 물 분자를 떼어내는 탈수화 과정을 거친다. 일부 연구에서는 중심 공동 내에 이미 존재하는 다른 칼륨 이온들의 전기적 반발력이 새로 들어오는 이온을 밀어내어, 전체적인 이온 통과 속도(처리량)를 높이는 데 기여할 수 있다고 본다.
5. 조절 (Regulation)
칼륨 통로 구멍을 통한 이온의 흐름은 게이팅과 불활성화라는 두 가지 관련 프로세스에 의해 조절된다. 게이팅은 자극에 반응하여 통로를 열거나 닫는 것이고, 불활성화는 열린 칼륨 통로에서 전류가 빠르게 중단되고 통로가 다시 전도할 수 있는 능력이 억제되는 것이다. 두 과정 모두 통로 전도성을 조절하는 데 기여하지만, 각 과정은 여러 메커니즘에 의해 매개될 수 있다.
일반적으로 게이팅은 자극을 감지하고 차례로 통로 구멍을 여는 추가적인 구조 도메인에 의해 매개되는 것으로 생각된다. 이러한 도메인에는 BK 통로의 RCK 도메인[69][70][71]과 전압 개폐 K+ 통로의 전압 센서 도메인이 포함된다. 이러한 도메인은 자극에 반응하여 구멍 도메인의 세포 내 게이트(헬릭스 게이트)를 물리적으로 열어 칼륨 이온이 막을 통과하도록 하는 것으로 생각된다. 일부 통로에는 자극에 대한 반응을 조절하는 여러 조절 도메인 또는 부속 단백질이 있다. 메커니즘에 대한 논쟁이 계속되고 있지만, 원핵생물[72][73][74] 및 진핵생물[69][70][71] 통로의 RCK 도메인, KcsA의 pH 게이팅 도메인,[75] 순환 뉴클레오타이드 게이팅 도메인,[76] 및 전압 개폐 칼륨 통로를 포함하여 이러한 조절 도메인의 구조가 알려져 있다.[77][78]
N형 불활성화는 일반적으로 더 빠른 불활성화 메커니즘이며, 이는 "볼 앤 체인" 모델이라고 한다.[79] N형 불활성화는 통로의 N-말단 또는 관련된 단백질과의 상호 작용을 포함하며, 이는 구멍 도메인과 상호 작용하여 "볼"처럼 이온 전도 경로를 막는다. 또는 C형 불활성화는 선택적 필터 자체 내에서 발생한다고 생각되며, 필터 내의 구조적 변화로 인해 비전도성이 된다. C형 불활성화된 K+ 통로 필터에 대한 여러 구조 모델이 있지만[80][81][82] 정확한 메커니즘은 여전히 불분명하다.
6. 약리학 (Pharmacology)
칼륨 통로는 인체의 다양한 생리 기능 조절에 핵심적인 역할을 하므로, 중요한 약리학적 표적이 된다.[5] 칼륨 통로의 기능을 조절하는 물질들은 크게 통로를 통한 칼륨 이온(K+)의 흐름을 막는 칼륨 통로 차단제와 통로의 활성을 증가시키는 칼륨 통로 활성제로 나눌 수 있다. 이러한 약물들은 고혈압, 부정맥, 뇌전증, 제2형 당뇨병, 통증 등 다양한 질환의 치료 가능성을 보여주며 활발히 연구되고 있다.[94] 특정 칼륨 통로에만 선택적으로 작용하는 약물을 개발하는 것은 여전히 어려운 과제이며, 표적 외 효과로 인한 부작용 발생 가능성도 고려해야 한다.[93]
6. 1. 차단제 (Blockers)
칼륨 통로 차단제는 통로를 통한 칼륨 이온(K+)의 흐름을 억제하는 물질이다. 이들은 선택성 필터 내에서 칼륨 결합과 경쟁하거나, 필터 외부에서 결합하여 이온 전도를 막는 방식으로 작용한다.대표적인 차단제 중 하나는 4급 암모늄 이온으로, 세포 외부 표면[83][84] 또는 통로의 중심 공동에 결합한다.[85] 특히 중심 공동에서의 차단을 위해서는 통로가 열려 있어야 하므로, 4급 암모늄 이온은 '열린 통로 차단제'로도 알려져 있다.[86]
바륨 이온(Ba2+) 또한 선택성 필터 내에서 높은 친화도로 결합하여 칼륨 통로 전류를 강력하게 차단할 수 있다.[87][88][89][90][91][92] 이러한 작용은 흥분성 세포의 칼륨 통로 활성을 억제하며, 바륨 독성의 주요 원인으로 지목된다.
덴드로톡신과 같은 일부 독소 역시 칼륨 통로를 차단하는 기전을 통해 강력한 독성을 나타낸다.[5]
의학적으로는 4-아미노피리딘 및 3,4-디아미노피리딘과 같은 칼륨 통로 차단제가 다발성 경화증과 같은 질환의 치료를 위해 연구되고 있다.[93] 그러나 이러한 약물은 표적 외 효과로 인해 부작용을 일으킬 수 있다. 특히 심장의 hERG 칼륨 통로에 작용하여 QT 연장 증후군을 유발할 가능성이 있으며, 이는 잠재적으로 생명을 위협할 수 있는 심각한 상태이다. 따라서 모든 신약은 개발 단계에서 심장 안전성 확보를 위해 임상 전 테스트를 거치게 된다.
6. 2. 활성제 (Activators)
칼륨 통로 활성제는 칼륨 통로의 활성을 증가시키는 물질로, 칼륨 통로 개폐기의 일종이다.7. 무스카린 칼륨 통로 (Muscarinic potassium channel)
일부 칼륨 통로는 무스카린 수용체에 의해 활성화되며, 이를 무스카린 칼륨 통로(IKACh)라고 부른다. 이 통로는 G 단백질 결합 내향 정류 칼륨 통로(GIRK)의 한 종류로, 4가지 하위 유형(GIRK1~GIRK4) 중 2개의 GIRK1과 2개의 GIRK4 소단위체가 결합한 이종사량체 구조를 가진다.[94][95]
대표적인 예로 심장에 존재하는 무스카린 칼륨 통로를 들 수 있다. 부교감 신경의 신호가 M2 무스카린 수용체를 활성화시키면, 이 통로를 통해 칼륨 이온이 세포 밖으로 나가는 전류가 발생하여 심박수를 느리게 조절한다.[96][97]
8. 예술 작품
로데릭 매키넌은 KcsA 칼륨 통로를 기반으로 한 약 1.52m 높이의 조각품인 ''아이디어의 탄생''을 의뢰했다.[98] 이 작품은 통로의 내부를 나타내는 철사 물체와 통로 구조의 주요 공동을 나타내는 불어진 유리 물체를 포함하고 있다.
참조
[1]
논문
Ion channels and synaptic organization: analysis of the Drosophila genome
2000-04
[2]
서적
Ion channels of excitable membranes
Sinauer
[3]
서적
Principles of Neural Science
McGraw-Hill
[4]
서적
The Alkali Metal Ions: Their Role for Life
Springer
2016
[5]
논문
Purification and subunit structure of a putative K+-channel protein identified by its binding properties for dendrotoxin I
1988-07
[6]
서적
Pharmacology
Churchill Livingstone
[7]
논문
Electrostatic interaction between charybdotoxin and a tetrameric mutant of Shaker K(+) channels
2000-05
[8]
논문
A strongly interacting pair of residues on the contact surface of charybdotoxin and a Shaker K+ channel
1996-01
[9]
논문
Peptide toxins and small-molecule blockers of BK channels
2016-01
[10]
논문
Mode of action of iberiotoxin, a potent blocker of the large conductance Ca(2+)-activated K+ channel
1992-08
[11]
논문
An apamin-sensitive Ca2+-activated K+ current in hippocampal pyramidal neurons
1999-04
[12]
논문
GAL-021, a new intravenous BKCa-channel blocker, is well tolerated and stimulates ventilation in healthy volunteers
2014-11
[13]
논문
Modulation of BK Channels by Ethanol
[14]
서적
Handbook of inorganic chemicals
https://archive.org/[...]
McGraw-Hill
2003
[15]
논문
Regulation of ROMK by extracellular cations
2001-02
[16]
논문
The inward rectifier current (IK1) controls cardiac excitability and is involved in arrhythmogenesis
2005-03
[17]
논문
Cardiac and renal inward rectifier potassium channel pharmacology: emerging tools for integrative physiology and therapeutics
2014-04
[18]
논문
Class III antiarrhythmic drug dronedarone inhibits cardiac inwardly rectifying Kir2.1 channels through binding at residue E224
2014-12
[19]
논문
Dual Mechanism for Inhibition of Inwardly Rectifying Kir2.x Channels by Quinidine Involving Direct Pore Block and PIP2-interference
2017-05
[20]
논문
Flecainide increases Kir2.1 currents by interacting with cysteine 311, decreasing the polyamine-induced rectification
2010-08
[21]
논문
Inhibition of G protein-activated inwardly rectifying K+ channels by ifenprodil
2006-03
[22]
논문
Centrally acting non-narcotic antitussives prevent hyperactivity in mice: Involvement of GIRK channels
2016-05
[23]
논문
"[Is the GIRK channel a possible target in the development of a novel therapeutic drug of urinary disturbance?]"
2011-04
[24]
논문
"[Novel antidepressant-like action of drugs possessing GIRK channel blocking action in rats]"
2010-05
[25]
논문
A novel high-affinity inhibitor for inward-rectifier K+ channels
1998-09
[26]
논문
The centrally acting non-narcotic antitussive tipepidine produces antidepressant-like effect in the forced swimming test in rats
https://kumadai.repo[...]
2009-12
[27]
논문
ML297 (VU0456810), the first potent and selective activator of the GIRK potassium channel, displays antiepileptic properties in mice
2013-09
[28]
논문
Glibenclamide, a blocker of K+(ATP) channels, shows antileishmanial activity in experimental murine cutaneous leishmaniasis
2006-12
[29]
논문
Gliclazide produces high-affinity block of KATP channels in mouse isolated pancreatic beta cells but not rat heart or arterial smooth muscle cells
2001-08
[30]
논문
Molecular background of leak K+ currents: two-pore domain potassium channels
http://repo.lib.semm[...]
2010-04
[31]
논문
Biophysical, pharmacological, and functional characteristics of cloned and native mammalian two-pore domain K+ channels
[32]
논문
A neuronal two P domain K+ channel stimulated by arachidonic acid and polyunsaturated fatty acids
1998-06
[33]
논문
Potassium leak channels and the KCNK family of two-P-domain subunits
https://escholarship[...]
2001-03
[34]
논문
A novel two-pore domain K+ channel, TRESK, is localized in the spinal cord
2003-07
[35]
논문
The two-pore domain K+ channel, TRESK, is activated by the cytoplasmic calcium signal through calcineurin
2004-04
[36]
논문
Local anesthetic inhibition of baseline potassium channels with two pore domains in tandem
1999-04
[37]
논문
Functional characterisation of human TASK-3, an acid-sensitive two-pore domain potassium channel
2001-03
[38]
논문
Amide local anesthetics potently inhibit the human tandem pore domain background K+ channel TASK-2 (KCNK5)
2003-07
[39]
논문
Inhibition of human TREK-1 channels by bupivacaine
2003-06
[40]
논문
TWIK-1, a ubiquitous human weakly inward rectifying K+ channel with a novel structure
1996-03
[41]
논문
TASK, a human background K+ channel to sense external pH variations near physiological pH
1997-09
[42]
논문
Cloning and expression of a novel pH-sensitive two pore domain K+ channel from human kidney
1998-11
[43]
논문
Cloning, localisation and functional expression of the human orthologue of the TREK-1 potassium channel
2000-04
[44]
웹사이트
UniProtKB - Q9NPC2 (KCNK9_HUMAN)
https://www.uniprot.[...]
Uniprot
2019-05-29
[45]
논문
Inhibition of the human two-pore domain potassium channel, TREK-1, by fluoxetine and its metabolite norfluoxetine
2005-03
[46]
논문
Inhalational anesthetics activate two-pore-domain background K+ channels
1999-05
[47]
논문
Volatile anesthetics activate the human tandem pore domain baseline K+ channel KCNK5
2000-06
[48]
논문
3,4-diaminopyridine. A potent new potassium channel blocker
1978-06
[49]
논문
Selective inhibition of K(+)-stimulation of Na,K-ATPase by bretylium
1991-12
[50]
논문
The selective inhibition of delayed potassium currents in nerve by tetraethylammonium ion
1967-05
[51]
논문
Interaction of tetraethylammonium ion derivatives with the potassium channels of giant axons
1971-10
[52]
웹사이트
Amiodarone
https://www.drugbank[...]
Drugbank
2019-05-28
[53]
논문
New molecular targets for antiepileptic drugs: alpha(2)delta, SV2A, and K(v)7/KCNQ/M potassium channels
2008-07
[54]
논문
The structure of the potassium channel: molecular basis of K+ conduction and selectivity
1998-04
[55]
논문
Structural conservation in prokaryotic and eukaryotic potassium channels
1998-04
[56]
논문
The vision of the pore
1998-04
[57]
웹사이트
The Nobel Prize in Chemistry 2003
http://nobelprize.or[...]
The Nobel Foundation
2007-11-16
[58]
논문
Chemistry of ion coordination and hydration revealed by a K+ channel-Fab complex at 2.0 A resolution
2001-11
[59]
서적
Molecular Cell Biology
W. H. Freeman and Company
2016
[60]
논문
Protein interactions central to stabilizing the K+ channel selectivity filter in a four-sited configuration for selective K+ permeation
2011-10
[61]
논문
Importance of hydration and dynamics on the selectivity of the KcsA and NaK channels
2007-02
[62]
논문
Control of ion selectivity in potassium channels by electrostatic and dynamic properties of carbonyl ligands
2004-10
[63]
논문
Tuning ion coordination architectures to enable selective partitioning
2007-08
[64]
논문
The predominant role of coordination number in potassium channel selectivity
2007-10
[65]
논문
Selectivity in K+ channels is due to topological control of the permeant ion's coordinated state
2007-05
[66]
논문
Tuning the ion selectivity of tetrameric cation channels by changing the number of ion binding sites
2011-01
[67]
논문
A comparison between two prokaryotic potassium channels (KirBac1.1 and KcsA) in a molecular dynamics (MD) simulation study
2006-03
[68]
논문
Energetic optimization of ion conduction rate by the K+ selectivity filter
2001-11
[69]
논문
Structure of the human BK channel Ca2+-activation apparatus at 3.0 A resolution
2010-07
[70]
논문
Structure of the gating ring from the human large-conductance Ca(2+)-gated K(+) channel
2010-07
[71]
논문
Structure of the RCK domain from the E. coli K+ channel and demonstration of its presence in the human BK channel
2001-03
[72]
논문
Crystal structure and mechanism of a calcium-gated potassium channel
2002-05
[73]
논문
Distinct gating mechanisms revealed by the structures of a multi-ligand gated K(+) channel
2012-12
[74]
논문
Crystal structure of a potassium ion transporter, TrkH
2011-03
[75]
논문
Mechanism of activation gating in the full-length KcsA K+ channel
2011-07
[76]
논문
Structural basis of ligand activation in a cyclic nucleotide regulated potassium channel
2004-11
[77]
논문
X-ray structure of a voltage-dependent K+ channel
2003-05
[78]
논문
Crystal structure of a mammalian voltage-dependent Shaker family K+ channel
2005-08
[79]
논문
Fast Inactivation of Voltage-Gated K(+) Channels: From Cartoon to Structure
1998-08
[80]
논문
Mechanism for selectivity-inactivation coupling in KcsA potassium channels
2011-03
[81]
논문
Structural mechanism of C-type inactivation in K(+) channels
2010-07
[82]
논문
Structural basis for the coupling between activation and inactivation gates in K(+) channels
2010-07
[83]
논문
Ions and blockers in potassium channels: insights from free energy simulations
2005-02
[84]
논문
Structure-activity relationship for extracellular block of K+ channels by tetraalkylammonium ions
2003-11
[85]
논문
The voltage-dependent gate in MthK potassium channels is located at the selectivity filter
2013-02
[86]
논문
The internal quaternary ammonium receptor site of Shaker potassium channels
1993-03
[87]
논문
Potassium-selective block of barium permeation through single KcsA channels
2011-10
[88]
논문
Potassium blocks barium permeation through a calcium-activated potassium channel
1988-11
[89]
논문
Structural and thermodynamic properties of selective ion binding in a K+ channel
2007-05
[90]
논문
The barium site in a potassium channel by x-ray crystallography
2000-03
[91]
논문
The conserved potassium channel filter can have distinct ion binding profiles: structural analysis of rubidium, cesium, and barium binding in NaK2K
2014-08
[92]
논문
Ionic interactions of Ba2+ blockades in the MthK K+ channel
2014-08
[93]
논문
Potassium channel blockers in multiple sclerosis: neuronal Kv channels and effects of symptomatic treatment
2006-07
[94]
논문
The G-protein-gated atrial K+ channel IKACh is a heteromultimer of two inwardly rectifying K(+)-channel proteins
1995-03
[95]
논문
Number and stoichiometry of subunits in the native atrial G-protein-gated K+ channel, IKACh
1998-02
[96]
논문
Identification of domains conferring G protein regulation on inward rectifier potassium channels
1995-11
[97]
논문
Structure, G protein activation, and functional relevance of the cardiac G protein-gated K+ channel, IKACh
http://www.annalsnya[...]
1999-04
[98]
간행물
The crucible: Art inspired by science should be more than just a pretty picture
http://www.rsc.org/c[...]
2008-03
[99]
서적
Ion channels of excitable membranes
Sinauer
[100]
서적
[[:en:Principles of Neural Science|Principles of Neural Science]]
McGraw-Hill
[101]
문서
(訳注)直列ポアドメインは「Tandem pore domain」に対する適切な日本語が見つからなかったため、暫定的に名付けた。
[102]
서적
Pharmacology
Churchill Livingstone
[103]
논문
The structure of the potassium channel: molecular basis of K+ conduction and selectivity
[104]
논문
Chemistry of ion coordination and hydration revealed by a K+ channel-Fab complex at 2.0 Â resolution
[105]
논문
Ion channels and synaptic organization: analysis of the Drosophila genome
2000-04
[106]
서적
Ion channels of excitable membranes
https://archive.org/[...]
Sinauer
[107]
서적
Principles of Neural Science
McGraw-Hill
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com